este capítulo es más relevante para la sección F8 (ii) del Plan de estudios primario del CICM 2017, que espera que los candidatos al examen puedan «describir el transporte de dióxido de carbono en la sangre, incluido el efecto Haldano y el cambio de cloruro»., A los examinadores universitarios les encanta este tema, y ha surgido varias veces en los artículos de la primera parte:
- Pregunta 1 del primer artículo de 2020
- Pregunta 5 del segundo artículo de 2018
- Pregunta 13 del primer artículo de 2015
- Pregunta 1 del segundo artículo de 2012
- Pregunta 6 del primer artículo de 2012
de los comentarios del examinador universitario para estas preguntas, ninguno es más útil que los comentarios para la pregunta 13 del primer documento de 2015, que ofrecen un esquema claro de lo que se esperaba.,
En resumen:
el CO2 es transportado por tres mecanismos principales:
- como bicarbonato (HCO3- ), 70-90% del contenido total de CO2 en sangre
- combinado con agua, el CO2 forma ácido carbónico, que a su vez forma bicarbonato:
CO2 + H2O H H2CO3 h HCO3- + h+- esto sucede principalmente en los glóbulos rojos
- El aumento de HCO3 intracelular – conduce al intercambio de bicarbonato y cloruro, el cambio de cloruro. El cloruro es absorbido por el RBCSs y el bicarbonato es liberado.,
- Por lo tanto, la concentración de cloruro es menor en la sangre venosa sistémica que en la sangre arterial sistémica
- Como carbamatos, las bases conjugadas de ácido carbamino (aproximadamente 10-20%)
- Bases conjugadas disociadas de ácidos carbamino, que se forman en la reacción espontánea de R-NH2 y CO2.,
- Las reservas de carbamino intracelulares (RBC) son las mayores: la hemoglobina, particularmente la hemoglobina desoxigenada, tiene una alta afinidad por el CO2, mientras que la mayoría de las otras proteínas no
- Como gas de CO2 disuelto, aproximadamente el 10%
- La Ley de Henry establece que la cantidad de gas disuelto en un líquido es proporcional a su presión parcial por encima del líquido
- Por lo tanto, por cada 1 mmHg de pCO2 la concentración en sangre aumenta en aproximadamente 0.,03 mmol/l
- Por lo tanto, el CO2 es 10-20 veces más soluble que el oxígeno
- ácido carbónico:
- Existe una proporción minúscula de dióxido de carbono total en esta forma, es decir, no es una contribución importante al transporte de CO2
hay una diferencia entre el contenido de CO2 arterial y venoso:
- la sangre tiene un contenido total de CO2 de aproximadamente 22,5 mmol/L
(o 520 ml/l)- la sangre arterial tiene un contenido total de CO2 de aproximadamente 20.,5 mmol/l
(O 480ml/L)- gran parte de esta diferencia se debe al aumento de la concentración de bicarbonato (85%)
- parte de esta diferencia también se debe al efecto Haldane:
- La Desoxihemoglobina tiene aproximadamente 3,5 veces la afinidad por el CO2 en comparación con la oxihemoglobina
- Esto aumenta la capacidad de unión al CO2 de la sangre venosa
- La Desoxihemoglobina también es un mejor buffer que la oxihemoglobina, lo que aumenta la capacidad de los glóbulos rojos para transportar HCO3-
las referencias más relevantes para este tema son el capítulo 4.,5 de «Acid-Base Physiology» de Brandis, así como este excelente artículo de revisión gratuito de Geers & Gross (2000). Si uno es capaz de hacerlo, uno debe hacer todo lo posible para obtener acceso a Klocke (2011), que desafortunadamente está pagado, pero que es integral más allá de todo reproche y mejor referenciado que el de Nunn.
proporciones de diferentes mecanismos de transporte de CO2
parece haber cierto desacuerdo entre las publicaciones con respecto a cuánto CO2 se transporta y dónde., Por ejemplo, varios artículos de revisión de nivel básico (como Arthurs & Sudhakar, 2005) afirman con autoridad que «aproximadamente el 75% del dióxido de carbono es transporte (sic) en los glóbulos rojos y el 25% en el plasma». En contraste, Geers & Gross (2000, Tabla 1) da un conjunto bastante diferente de números, casi totalmente al revés.,ood referencias y el primero no, el autor le había atribuido crédulamente una mayor credibilidad y crudamente reensamblado sus datos prestados como una serie de cilindros de color:
![]()
o, para presentarlo de una manera ligeramente diferente:
![]()
o, si se prefiere datos reales:
| sangre arterial (mmol/l) |
sangre venosa mixta (mmol)l) |
|
| 0.,68 | 0.78 | |
| Plasma: bicarbonate | 13.52 | 14.51 |
| Plasma: carbamate | 0.3 | 0.3 |
| RBC dissolved CO2 | 0.4 | 0.46 |
| RBC bicarbonate | 5.01 | 5.46 |
| RBC carbamate | 0.75 | 0.84 |
| Total CO2 content of acellular plasma | 14.5 | 15.59 |
| Total CO2 content of RBCs | 6.16 | 6.,76 |
Este es un útil segue en la discusión de la diferencia entre el contenido de dióxido de caron arterial y venoso.
diferencia entre el contenido de CO2 arterial y venoso
aunque esto no se indica explícitamente en la pregunta stem («describir el transporte de dióxido de Carbono (CO2) en la sangre»), los examinadores universitarios esperaban que los candidatos al examen produjeran «una tabla de la contribución de cada forma de transporte al contenido arterial y venoso y la diferencia AV» en su respuesta a la Pregunta 5 del segundo artículo de 2018.,
la diferencia normal entre PaCO2 y PvCO2 generalmente se dice que es de aproximadamente 2-6 mmHg (Groeneveld, 1998), pero esta es una diferencia de tensión más que de contenido. El contenido realmente debe medirse en unidades de contenido, por ejemplo. Algunos autores reportan ml/L, y otros mmol/L. Los datos de Geers & Gross (2000) sugieren que esta diferencia no es masiva.
de todos modos. En el torrente sanguíneo arterial, los complejos de carbamato CO2-proteína y el CO2 disuelto generalmente representan menos del 15% de la capacidad total de transporte de CO2 en plasma. El resto se compone de HCO3 -., Dentro de los eritrocitos, donde la concentración de hemoglobina es de alrededor de 330 g / L, la cantidad de compuestos de carbamato es mayor, pero el bicarbonato sigue siendo el jugador principal. A medida que el ejercicio aumenta la cantidad de CO2 producido por los tejidos, la proporción de gas CO2 disuelto en plasma y eritrocitos aumenta (¡triples!) pero este aumento en el gas disuelto tiene un efecto menor en el contenido total de CO2 en sangre.
En resumen, en la sangre humana el CO2 se transporta principalmente al convertirse en bicarbonato., Una manera aún más efectiva de representar esto se puede tomar prestada de la fisiología respiratoria de Nunn. En el Capítulo 10 de la 8a edición (que trata sobre el transporte de dióxido de carbono) existe un diagrama que ilustra el cambio en el transporte de CO2 como se representa contra la presión parcial de CO2. También es probablemente el diagrama solicitado en los comentarios del colegio para la pregunta 13 del primer artículo de 2015, donde los examinadores mencionaron que «un diagrama de contenido de pCO2 v CO2 fue útil»., Se parece mucho a esto:
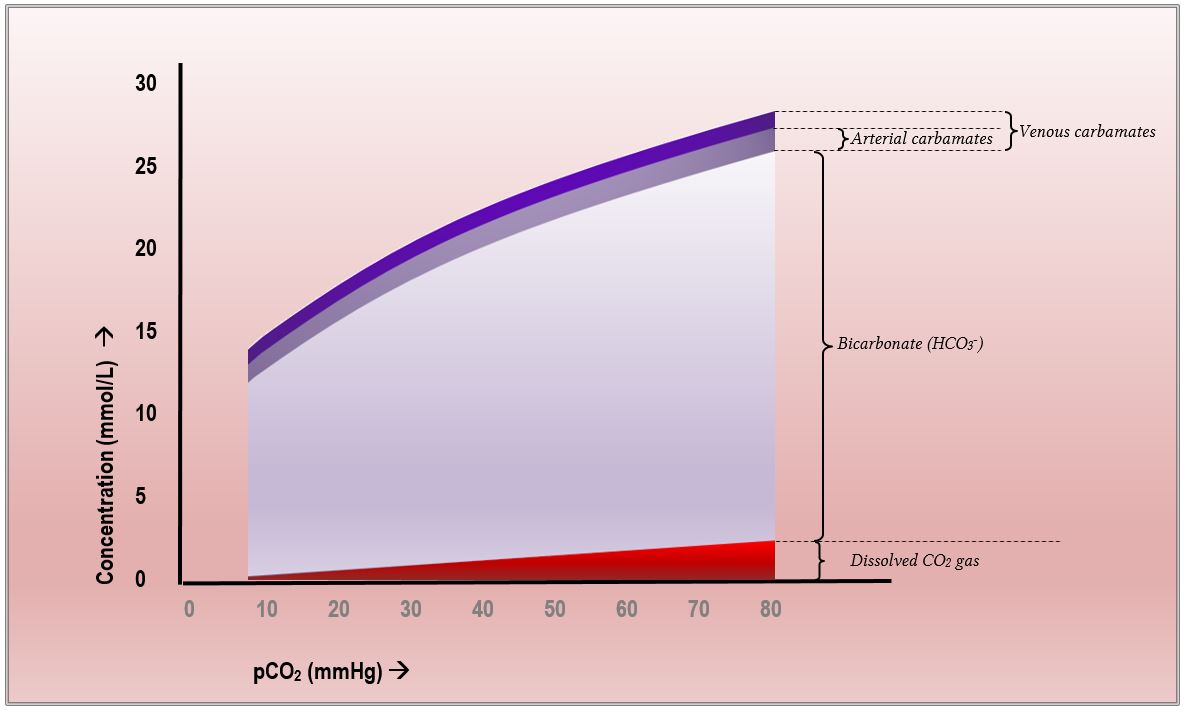
Esta es de hecho la curva de disociación de CO2, similar a la curva de disociación oxígeno-hemoglobina pero distinta porque a diferencia del oxígeno, el CO2 se transporta de tres maneras diferentes. Como se puede ver claramente, con el aumento de pCO2 la concentración de bicarbonato cambia más, el CO2 disuelto aumenta modestamente de manera lineal, y el contenido de carbamato apenas cambia en absoluto.
transporte de CO2 como gas disuelto
el CO2 pasa a ser uno de los gases que disfrutan de una solubilidad en agua relativamente buena., Por cada 1 mmHg de CO2, aproximadamente 0,03 mmol/L entra en solución. Esta relación es razonablemente lineal sobre un rango de supervivencia de presiones y temperaturas.

en solución, el CO2 se combinará reversiblemente con el agua para formar ácido carbónico, pero lo hace con gran reticencia. Uno puede describir esta renuencia matemáticamente, y expresarla como una constante de equilibrio de hidratación (constante de Henry). Esta constante se deriva dividiendo la concentración de H2CO3 por la concentración de CO2 acuoso. Por lo tanto, a 25°C, la constante de Henry para el CO2 es solo alrededor de 1.,7 x10-3, lo que significa que en cualquier frasco de agua, la mayoría del CO2 disuelto permanece como gas disuelto, y solo una pequeña fracción (0.17%) está presente como H2CO3.
de todos modos. El contenido total de CO2 disuelto de la sangre se puede calcular a partir de la relación anterior ( 0.03 mmol/l/mmHg); por lo tanto, es 1.2 mmol/L.
transporte de CO2 como bicarbonato
conversión de CO2 en ácido carbónico y bicarbonato
la combinación de CO2 y H2O conduce a la formación de H2CO3 de una manera totalmente espontánea sin catalizar., El CO2 actúa como un ácido de Lewis en este entorno; al no tener protones para donarlo, actúa como «algo capaz de formar un enlace covalente con un par de electrones».
la reacción espontánea con el agua es en realidad bastante lenta. Para cantidades clínicamente significativas de CO2, la reacción de disociación sin catalizar CO2 + H2O H H2CO3 puede tardar aproximadamente 3-4 segundos en equilibrarse a la mitad, y hasta un minuto en alcanzar el equilibrio completo (y eso es en la temperatura corporal de soporte de la reacción de vapor, 37°C). Obviamente eso es demasiado lento., No habría tiempo suficiente para que esta reacción llegara al equilibrio durante el rápido tránsito de glóbulos rojos a través de los capilares pulmonares de intercambio de gases, y no se produciría la eliminación de CO2.

afortunadamente, el interior de los eritrocitos (así como las paredes de los vasos sanguíneos) están llenos de anhidrasa carbónica (específicamente CA Tipo II) , que es quizás la enzima de acción más rápida en el cuerpo humano, y con la ayuda de la cual esta reacción progresa 25,000 veces más rápido, alcanzando el 95% de finalización en 2 milisegundos., En las macroescalas de ventilación, medidas en segundos y minutos, esta reacción puede considerarse esencialmente instantánea. Debido a este tipo de soporte catalítico, la conversión de HCO3 – y H+ de nuevo en CO2 y H2O puede ocurrir en los capilares pulmonares durante el tiempo de tránsito capilar pulmonar muy breve.
de hecho, la anhidrasa carbónica es tan eficiente que se requieren relativamente pocas moléculas enzimáticas para mantener un nivel completamente normal de intercambio de CO2. Parecería que uno requiere quizás tan poco como 2% de enzima desbloqueada para continuar., Considere que este sistema enzimático es bloqueado rutinariamente por razones terapéuticas por medicamentos como la acetazolamida. De hecho, las sobredosis masivas de inhibidores de la anhidrasa carbónica han sido sorprendentemente sobrevivibles, a pesar de la acidosis metabólica severa que ocasionalmente se produce.
transporte de CO2 como compuestos de carbamato
Los compuestos de carbamato son las bases conjugadas disociadas de los ácidos de carbamino, que se forman en la reacción espontánea de R-NH2 y CO2.,

por lo tanto, un «carbamato» es una base conjugada que actúa como un dispositivo de almacenamiento de CO2, y puede contribuir hasta el 14% de la capacidad total de carga de CO2 de la sangre. La etapa de Transición «carbamino» descrita anteriormente es-como el ácido carbónico – una figura del habla en lugar de un miembro macroscópico apropiado del torrente sanguíneo., A pH fisiológico se disocia en carbamato y H + tan rápido y tan completamente que su concentración es insignificante, y para todos los efectos puede ser
el contenido de carbamato del plasma es bastante pobre porque las proteínas séricas (por ejemplo. globulinas) tienen poco interés en el transporte de CO2, y tienden a formar carbamatos con la mayor reticencia. El CO2 realmente solo puede unirse a las cadenas laterales de lisina y arginina, y cualquier proteína plasmática aleatoria solo puede tener una de estas cadenas laterales expuesta y disponible para la Unión del CO2., Hay más sitios de unión disponibles dentro de los glóbulos rojos, donde la hemoglobina (especialmente la desoxihemoglobina) está disponible en una concentración alta.
esta escasez de sitios de unión podría hacer pensar que los carbamatos no juegan un papel importante en el transporte de CO2 en general, y específicamente en su transporte en plasma. De hecho, los carbamatos contribuyen relativamente poco a la capacidad general de carga de CO2 de la sangre humana. Su importancia se vuelve mucho mayor si por alguna razón la mayor parte de su anhidrasa carbónica está desactivada., En ese caso, de repente este sistema de transporte olvidado se vuelve muy relevante, ya que puede funcionar con CO2 bruto y no requiere H2CO3 como intermediario. Además, el sistema carbamato hace la mayor contribución a la diferencia entre la concentración arterial y venosa de CO2, principalmente debido al efecto Haldano que se discute en otra parte.,
factores que influyen en la capacidad de carga de CO2 de las proteínas
el transporte de CO2 como carbamatos está influenciado por los siguientes factores:
- oxigenación de hemoglobina (más desoxigenada= mayor capacidad de CO2)
- pH en el entorno local (menor pH = mayor capacidad de CO2)
dentro de los glóbulos rojos, el CO2 se une fácilmente a las cadenas laterales de aminoácidos de la molécula de hemoglobina, particularmente cuando se desoxigena. De hecho, la desoxihemoglobina tiene aproximadamente 3.,5 veces la afinidad por el CO2 en comparación con la oxihemoglobina; este aumento inducido por la hipoxia en la capacidad de carga de CO2 se denomina efecto Haldane. Esto es exactamente lo que quieres en el tejido de ejercicio, que acaba de consumir todo el oxígeno en su volumen de sangre local, y ha producido una gran cantidad de CO2. Por lo tanto, el contenido de carbamato de la sangre venosa podría potencialmente (a un SvO2 del 0%) ser aproximadamente el triple del contenido de carbamato de la sangre arterial. Por supuesto, en realidad, la diferencia a-v nunca es tan grande, porque la sangre venosa nunca es tan hipóxica.,
la presencia de grupos carbamato en regiones críticas de la molécula de hemoglobina tiende a estabilizar la forma desoxigenada, lo que disminuye la afinidad de la Unión hemoglobina-oxígeno. Este efecto (el efecto Bohr) se analiza con mayor detalle junto con los demás factores que afectan a la afinidad de la hemoglobina por el oxígeno. Baste decir que la presencia de una gran cantidad de CO2 disminuye esta afinidad y promueve la liberación de más oxígeno de la hemoglobina, que de nuevo es precisamente lo que se necesita en el ejercicio del tejido.




