LTP a fost cel mai bine studiat în hipocampul mamifer, o zonă a creierului care este deosebit de importantă în formarea și/sau recuperarea unor forme de memorie (vezi capitolul 31). La om, imagistica funcțională arată că hipocampul uman este activat în timpul anumitor tipuri de sarcini de memorie și că deteriorarea hipocampului duce la incapacitatea de a forma anumite tipuri de amintiri noi. La rozătoare, neuronii hipocampali declanșează potențiale de acțiune numai atunci când un animal se află în anumite locații., Astfel de „celule de loc” par să codifice amintirile spațiale, o interpretare susținută de faptul că deteriorarea hipocampală împiedică șobolanii să dezvolte competență în sarcinile de învățare spațială (figura 25.4). Deși multe alte zone ale creierului sunt implicate în procesul complex de formare, stocare și recuperare a memoriei, aceste observații au determinat mulți cercetători să studieze această formă particulară de plasticitate sinaptică în hipocampus.

figura 25.4
învățarea spațială la rozătoare., (A) șobolanii sunt plasați într-o arenă circulară (aproximativ dimensiunea și forma bazinului de vânătoare al unui copil) umplută cu apă tulbure. Arena în sine este featureless, dar mediul înconjurător conține indicii poziționale, cum ar fi windows, (mai mult…lucrul la LTP a început la începutul anilor 1970, când Timothy Bliss și colegii săi de la Mill Hill din Anglia au descoperit că câteva secunde de stimulare electrică de înaltă frecvență pot îmbunătăți transmisia sinaptică în hipocampul iepurelui zile sau chiar săptămâni., Mai recent, însă, progresul în înțelegerea mecanismului LTP s-a bazat foarte mult pe studiile in vitro ale feliilor de hipocampus viu. Aranjamentul neuronilor permite hipocampului să fie secționat astfel încât majoritatea circuitelor relevante să fie lăsate intacte. În astfel de preparate, corpurile celulare ale neuronilor piramidali se află într-un singur strat dens ambalat, care este ușor evident (figura 25.5). Acest strat este împărțit în mai multe regiuni distincte, cele mai importante fiind CA1 și CA3., „CA” se referă la Cornu Ammon, un nume Latin pentru cornul lui Ammon—cornul Berbecului care seamănă cu forma hipocampului. Dendritele celulelor piramidale din regiunea CA1 formează o bandă groasă (stratul radiatum), unde primesc sinapse de la colateralele Schaffer, axonii celulelor piramidale din regiunea CA3. O mare parte din lucrările privind LTP s-au concentrat pe conexiunile sinaptice dintre colateralele Schaffer și celulele piramidale CA1. Stimularea electrică a colateralelor Schaffer generează potențiale postsinaptice excitatorii (EPSPs) în celulele CA1 postsinaptice (figura 25.6)., Dacă colateralele Schaffer sunt stimulate doar de două sau trei ori pe minut, dimensiunea EPSP evocată în neuronii CA1 rămâne constantă. Cu toate acestea, un scurt tren de înaltă frecvență de stimuli către aceiași axoni provoacă LTP, ceea ce este evident ca o creștere de lungă durată a amplitudinii EPSP. LTP apare, de asemenea, la multe alte sinapse, atât în hipocampus, cât și într-o varietate de alte regiuni ale creierului, inclusiv cortexul, amigdala și cerebelul.

figura 25.,5
diagrama unei secțiuni prin hipocampul rozătoarelor care prezintă regiunile majore, căile excitatorii și conexiunile sinaptice. Potențarea pe termen lung a fost observată la fiecare dintre cele trei conexiuni sinaptice prezentate aici.
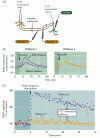
figura 25.6
potențarea pe termen lung a sinapselor colaterale Schaffer-CA1. (A) aranjament pentru înregistrarea transmisiei sinaptice; doi electrozi stimulatori (1 și 2) activează fiecare populații separate de colaterale Schaffer, asigurând astfel testarea și controlul sinaptic (mai mult…,LTP din sinapsa colaterală Schaffer prezintă mai multe proprietăți care o fac un mecanism neural atractiv pentru stocarea informațiilor. În primul rând, LTP este dependent de stat: gradul de depolarizare a celulei postsinaptice determină dacă apare sau nu LTP (figura 25.7). Dacă un singur stimul pentru colateralele Schaffer—care nu ar provoca în mod normal LTP—este asociat cu depolarizarea puternică a celulei CA1 postsinaptice, dimensiunea EPSP este mărită., Creșterea apare numai dacă activitățile asociate ale celulelor presinaptice și postsinaptice sunt strâns legate în timp, astfel încât depolarizarea postsinaptică puternică are loc în aproximativ 100 ms de eliberare a transmițătorului presinaptic. Reamintim că o cerință pentru care coincide activarea presinaptice și postsinaptică elemente este semnul distinctiv al Hebbian postulat (vezi Capitolul 24), o încercare de a stabili un cadru teoretic de sinaptice modificări de fond al sistemului de învățare și memorie.

figura 25.,7
asocierea activității presinaptice și postsinaptice determină LTP. Stimulii unici aplicați unei intrări sinaptice colaterale Schaffer evocă EPSPs în neuronul CA1 postsynaptic. Numai acești stimuli nu provoacă nicio schimbare a puterii sinaptice. Cu toate acestea, atunci când (mai mult…LTP prezintă, de asemenea, proprietatea specificității de intrare: atunci când LTP este indusă de stimularea unei sinapse, aceasta nu apare în alte sinapse inactive care intră în contact cu același neuron (vezi figura 25.6)., Astfel, LTP este specific intrării în sensul că este limitat la sinapsele activate, mai degrabă decât la toate sinapsele dintr-o anumită celulă (figura 25.8 a). Această caracteristică a LTP este în concordanță cu implicarea sa în formarea memoriei. Dacă activarea unui set de sinapse a dus la potențarea tuturor celorlalte sinapse—chiar și a celor inactive -, ar fi dificil să se îmbunătățească selectiv anumite seturi de intrări, așa cum se presupune că este necesar pentru învățare și memorie (vezi și caseta B).
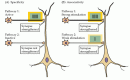
figura 25.,8
proprietățile LTP la un neuron piramidal CA1 care primește intrări sinaptice din două seturi independente de axoni colaterali Schaffer. (A) Activitatea puternică inițiază LTP la sinapsele active (calea 1) fără a iniția LTP la sinapsele inactive din apropiere (calea (mai mult…)
caseta B
sinapsele silențioase. o altă proprietate importantă a LTP este asociativitatea (figura 25.8 B). După cum sa menționat, stimularea slabă a unei căi nu va declanșa ea însăși LTP., Cu toate acestea, dacă o cale este slab activată în același timp în care o cale vecină pe aceeași celulă este puternic activată, ambele căi sinaptice suferă LTP. Această îmbunătățire selectivă a seturilor de intrări sinaptice activate în comun este adesea considerată un analog celular al condiționării asociative sau clasice. Mai general, asociativitatea este așteptată în orice rețea de neuroni care leagă un set de informații cu altul.,deși există în mod clar un decalaj între înțelegerea LTP a sinapselor hipocampale și înțelegerea plasticității comportamentale, această formă de plasticitate sinaptică oferă un mecanism neural plauzibil pentru schimbări de lungă durată într-o parte a creierului despre care se știe că este implicată în formarea anumitor tipuri de amintiri.




