LTP è stato studiato più a fondo nell’ippocampo dei mammiferi, un’area del cervello che è particolarmente importante nella formazione e/o nel recupero di alcune forme di memoria (vedi Capitolo 31). Negli esseri umani, l’imaging funzionale mostra che l’ippocampo umano viene attivato durante determinati tipi di attività di memoria e che il danno all’ippocampo provoca l’incapacità di formare determinati tipi di nuovi ricordi. Nei roditori, i neuroni ippocampali sparano potenziali d’azione solo quando un animale si trova in determinate posizioni., Tali” cellule del luogo ” sembrano codificare le memorie spaziali, un’interpretazione supportata dal fatto che il danno all’ippocampo impedisce ai ratti di sviluppare competenza nei compiti di apprendimento spaziale (Figura 25.4). Sebbene molte altre aree cerebrali siano coinvolte nel complesso processo di formazione, conservazione e recupero della memoria, queste osservazioni hanno portato molti ricercatori a studiare questa particolare forma di plasticità sinaptica nell’ippocampo.

Figura 25.4
Apprendimento spaziale nei roditori., (A) I ratti sono collocati in un’arena circolare (circa le dimensioni e la forma della piscina per bambini) piena di acqua torbida. L’arena stessa è senza caratteristiche, ma l’ambiente circostante contiene i segnali posizionali come windows, (altro…)
Il lavoro su LTP iniziò nei primi anni 1970, quando Timothy Bliss e i suoi colleghi di Mill Hill in Inghilterra scoprirono che alcuni secondi di stimolazione elettrica ad alta frequenza possono migliorare la trasmissione sinaptica nell’ippocampo del coniglio per giorni o addirittura settimane., Più recentemente, tuttavia, i progressi nella comprensione del meccanismo di LTP hanno fatto molto affidamento su studi in vitro su fette di ippocampo vivente. La disposizione dei neuroni consente all’ippocampo di essere sezionato in modo tale che la maggior parte dei circuiti rilevanti sia lasciata intatta. In tali preparazioni, i corpi cellulari dei neuroni piramidali si trovano in un unico strato densamente imballato che è facilmente evidente (Figura 25.5). Questo strato è diviso in diverse regioni distinte, le principali sono CA1 e CA3., ” CA ” si riferisce a Cornu Ammon, un nome latino per il corno di Ammon—il corno dell’ariete che ricorda la forma dell’ippocampo. I dendriti delle cellule piramidali nella regione CA1 formano una banda spessa (lo strato radiatum), dove ricevono sinapsi dai collaterali di Schaffer, gli assoni delle cellule piramidali nella regione CA3. Gran parte del lavoro su LTP si è concentrato sulle connessioni sinaptiche tra i collaterali di Schaffer e le cellule piramidali CA1. La stimolazione elettrica dei collaterali di Schaffer genera potenziali postsinaptici eccitatori (EPSP) nelle cellule CA1 postsinaptiche (Figura 25.6)., Se i collaterali di Schaffer vengono stimolati solo due o tre volte al minuto, la dimensione dell’EPSP evocato nei neuroni CA1 rimane costante. Tuttavia, un breve treno ad alta frequenza di stimoli agli stessi assoni causa LTP, che è evidente come un aumento duraturo dell’ampiezza EPSP. LTP si verifica anche in molte altre sinapsi, sia all’interno dell’ippocampo che in una varietà di altre regioni del cervello, tra cui la corteccia, l’amigdala e il cervelletto.

Figura 25.,5
Diagramma di una sezione attraverso l’ippocampo dei roditori che mostra le regioni principali, le vie eccitatorie e le connessioni sinaptiche. È stato osservato un potenziamento a lungo termine in ciascuna delle tre connessioni sinaptiche qui mostrate.
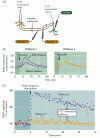
Figura 25.6
Potenziamento a lungo termine delle sinapsi collaterali di Schaffer-CA1. (A) Disposizione per la registrazione della trasmissione sinaptica; due elettrodi stimolanti (1 e 2) attivano ciascuno popolazioni separate di collaterali Schaffer, fornendo così test e controllo sinaptico (altro…,)
LTP della sinapsi collaterale di Schaffer presenta diverse proprietà che lo rendono un attraente meccanismo neurale per l’archiviazione delle informazioni. Innanzitutto, LTP è dipendente dallo stato: il grado di depolarizzazione della cellula postsinaptica determina se si verifica o meno LTP (Figura 25.7). Se un singolo stimolo ai collaterali di Schaffer-che normalmente non suscitano LTP-è accoppiato con una forte depolarizzazione della cellula CA1 postsinaptica, la dimensione dell’EPSP aumenta., L’aumento si verifica solo se le attività accoppiate delle cellule presinaptiche e postsinaptiche sono strettamente collegate nel tempo, in modo tale che la forte depolarizzazione postsinaptica si verifica entro circa 100 ms dal rilascio del trasmettitore presinaptico. Ricordiamo che un requisito per l’attivazione coincidente di elementi presinaptici e postsinaptici è il segno distintivo del postulato hebbiano (vedi Capitolo 24), un primo tentativo di stabilire un quadro teorico dei cambiamenti sinaptici alla base dell’apprendimento e della memoria.

Figura 25.,7
L’associazione dell’attività presinaptica e postsinaptica causa LTP. Singoli stimoli applicati a un input sinaptico collaterale di Schaffer evocano EPSP nel neurone CA1 postsinaptico. Questi stimoli da soli non provocano alcun cambiamento nella forza sinaptica. Tuttavia, quando il (più…)
LTP presenta anche la proprietà della specificità di input: quando LTP è indotto dalla stimolazione di una sinapsi, non si verifica in altre sinapsi inattive che contattano lo stesso neurone (vedi Figura 25.6)., Pertanto, LTP è specifico per l’input nel senso che è limitato alle sinapsi attivate piuttosto che a tutte le sinapsi su una data cella (Figura 25.8 A). Questa caratteristica di LTP è coerente con il suo coinvolgimento nella formazione della memoria. Se l’attivazione di un insieme di sinapsi ha portato a tutte le altre sinapsi—anche quelle inattive—potenziate, sarebbe difficile migliorare selettivamente determinati set di input, come presumibilmente richiesto per l’apprendimento e la memoria (vedi anche Riquadro B).
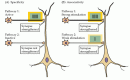
Figura 25.,8
Proprietà di LTP su un neurone piramidale CA1 che riceve input sinaptici da due insiemi indipendenti di assoni collaterali di Schaffer. (A) Una forte attività avvia LTP a sinapsi attive (pathway 1) senza avviare LTP a sinapsi inattive vicine (pathway (more…)
Casella B
Sinapsi silenziose.
Un’altra proprietà importante di LTP è l’associatività (Figura 25.8 B). Come notato, la debole stimolazione di un percorso non attiverà da sola LTP., Tuttavia, se una via è debolmente attivata nello stesso momento in cui una via vicina sulla stessa cellula è fortemente attivata, entrambe le vie sinaptiche subiscono LTP. Questo miglioramento selettivo di insiemi attivati congiuntamente di input sinaptici è spesso considerato un analogo cellulare del condizionamento associativo o classico. Più in generale, l’associatività è prevista in qualsiasi rete di neuroni che collega un insieme di informazioni con un altro.,
Sebbene vi sia chiaramente un divario tra la comprensione della LTP delle sinapsi ippocampali e la comprensione della plasticità comportamentale, questa forma di plasticità sinaptica fornisce un meccanismo neurale plausibile per cambiamenti duraturi in una parte del cervello che è nota per essere coinvolta nella formazione di determinati tipi di ricordi.




