Ce chapitre est le plus pertinent pour la Section F8(ii) du Programme primaire 2017 du CICM, qui s’attend à ce que les candidats à l’examen soient en mesure de « décrire le transport du dioxyde de carbone dans le sang, y compris l’effet Haldane, Les examinateurs du collège adorent ce sujet, et il est apparu plusieurs fois dans les articles de la première partie:
- Question 1 du premier article de 2020
- Question 5 du deuxième article de 2018
- Question 13 du premier article de 2015
- Question 1 du deuxième article de 2012
- Question 6 du premier article de 2012
parmi les commentaires des examinateurs de l’ordre pour ces questions, aucun n’est plus utile que les commentaires pour la question 13 du premier document de 2015, qui offrent un aperçu clair de ce qui était attendu.,
En résumé:
le CO2 est transporté par trois mécanismes majeurs:
- sous forme de bicarbonate (HCO3- ), 70-90% de la teneur en CO2 du sang total
- combiné à l’eau, le CO2 forme de l’acide carbonique, qui:
CO2 + H2O ⇌ H2CO3 ⇌ HCO3- + h+- cela se produit principalement dans les RBCs
- la montée en HCO3 – intracellulaire conduit à l’échange de bicarbonate et de chlorure, le déplacement du chlorure. Le chlorure est absorbé par les RBCSs et le bicarbonate est libéré.,
- ainsi, la concentration de chlorure est plus faible dans le sang veineux systémique que dans le sang artériel systémique
- sous forme de carbamates, les bases conjuguées de l’acide carbamino (environ 10-20%)
- Les bases conjuguées dissociées des carbamino-acides, qui se forment dans la réaction spontanée de R-NH2 et de CO2.,
- Les réserves de carbamino intracellulaires (RBC) sont les plus importantes: l’hémoglobine, en particulier l’hémoglobine désoxygénée, a une forte affinité pour le CO2, alors que la plupart des autres protéines ne le font pas
- En tant que gaz CO2 dissous, environ 10%
- La loi de Henry stipule que la quantité de gaz dissous dans un liquide est proportionnelle à sa pression partielle au-dessus du liquide
- ainsi, pour chaque 1 mmHg De pCO2, la concentration sanguine augmente d’environ 0.,03 mmol/L
- ainsi, le CO2 est 10 à 20 fois plus soluble que l’oxygène
- acide carbonique:
- une infime proportion de dioxyde de carbone total existe sous cette forme, c’est-à-dire qu’il n’est pas un contributr majeur au transport du CO2
le sang a une teneur totale en CO2 d’environ 22,5 mmol/l
(ou 520 ml/L)- le sang artériel a une teneur totale en CO2 d’environ 20.,5 mmol/l
(ou 480 ml/L)- Une grande partie de cette différence est due à l’augmentation de la concentration de bicarbonate (85%)
- Une partie de cette différence est également due à L’effet Haldane:
- La Désoxyhaémoglobine a environ 3,5 fois l’affinité pour le CO2 par rapport à l’oxyhaémoglobine
- cela augmente meilleur tampon que l’oxyhaémoglobine, ce qui augmente la capacité des RBCs à transporter HCO3 –
les références les plus pertinentes pour ce sujet sont le chapitre 4.,5 de « physiologie acido-basique » de Brandis, ainsi que cet excellent article gratuit de Geers & Gross (2000). Si l’on est capable de le faire, il faut tout faire pour avoir accès à Klocke (2011), qui est malheureusement payant, mais qui est complet sans reproche mieux référencé que Nunn.
Proportions des différents mécanismes de transport du CO2
Il semble y avoir un certain désaccord entre les publications quant à la quantité de CO2, Par exemple, divers articles de revue de base (tels que Arthurs & Sudhakar, 2005) indiquent avec autorité que « environ 75% du dioxyde de carbone est transporté (sic) dans les globules rouges et 25% dans le plasma ». En revanche, Geers & Gross (2000, Tableau 1) donne un ensemble de nombres assez différent, presque totalement inverse.,ood références et le premier ne le fait pas, l’auteur lui avait crédule attribué une plus grande crédibilité et a grossièrement remonté leurs données empruntées comme une série de cylindres colorés:
![]()
ou, pour les présenter d’une manière légèrement différente:
![]()
ou, si l’on préfère données réelles:
| sang artériel (mmol/l) |
sang veineux mixte (mmol)L) |
|
| plasma: dissous CO2 | 0.,68 | 0.78 |
| Plasma: bicarbonate | 13.52 | 14.51 |
| Plasma: carbamate | 0.3 | 0.3 |
| RBC dissolved CO2 | 0.4 | 0.46 |
| RBC bicarbonate | 5.01 | 5.46 |
| RBC carbamate | 0.75 | 0.84 |
| Total CO2 content of acellular plasma | 14.5 | 15.59 |
| Total CO2 content of RBCs | 6.16 | 6.,76 |
ceci est une séquence pratique dans la discussion de la différence entre la teneur en dioxyde de caron artériel et veineux.
différence entre la teneur en CO2 artériel et veineux
bien que cela ne soit pas explicitement indiqué dans la question stem (« décrire le transport de dioxyde de carbone (CO2) dans le sang »), les examinateurs du collège s’attendaient à ce que les candidats à l’examen produisent « un tableau de la contribution de chaque forme de transport au contenu artériel et veineux et à la différence AV » dans leur réponse à la Question 5 du deuxième article de 2018.,
la différence normale entre PaCO2 et PvCO2 est généralement d’environ 2-6 mmHg (Groeneveld, 1998), mais il s’agit d’une différence de tension plutôt que de contenu. Le contenu devrait vraiment être mesuré en unités de contenu, d’une part. Certains auteurs signalent ml/L, et d’autres mmol/L. Les données de Geers & Gross (2000) suggèrent que cette différence n’est pas massive.
de toute façon. Dans la circulation sanguine artérielle, les complexes carbamate CO2-protéine et le CO2 dissous représentent généralement moins de 15% de la capacité totale de transport de CO2 plasmatique. Le reste est composé de HCO3 -., À l’intérieur des érythrocytes, où le taux d’hémoglobine est d’environ 330g/L, la quantité de composés carbamates est la plus élevée, mais le bicarbonate reste le principal acteur. À mesure que l’exercice augmente la quantité de CO2 produite par les tissus, la proportion de gaz CO2 dissous dans le plasma et les érythrocytes augmente (Triple!) mais cette augmentation du gaz dissous a un effet mineur sur la teneur totale en CO2 dans le sang.
en bref, dans le sang total humain, le CO2 est transporté principalement en étant converti en bicarbonate., Un moyen encore plus efficace de représenter cela peut être emprunté à la physiologie respiratoire de Nunn. Dans le Chapitre 10 de la 8ème édition (qui concerne le transport du dioxyde de carbone), un diagramme existe qui illustre le changement du transport du CO2 en fonction de la pression partielle du CO2. C’est aussi probablement le diagramme demandé dans les commentaires du collège pour la Question 13 du premier document de 2015, où les examinateurs ont mentionné qu ‘ « un diagramme de la teneur en pCO2 v CO2 était utile »., Il ressemble beaucoup à ceci:
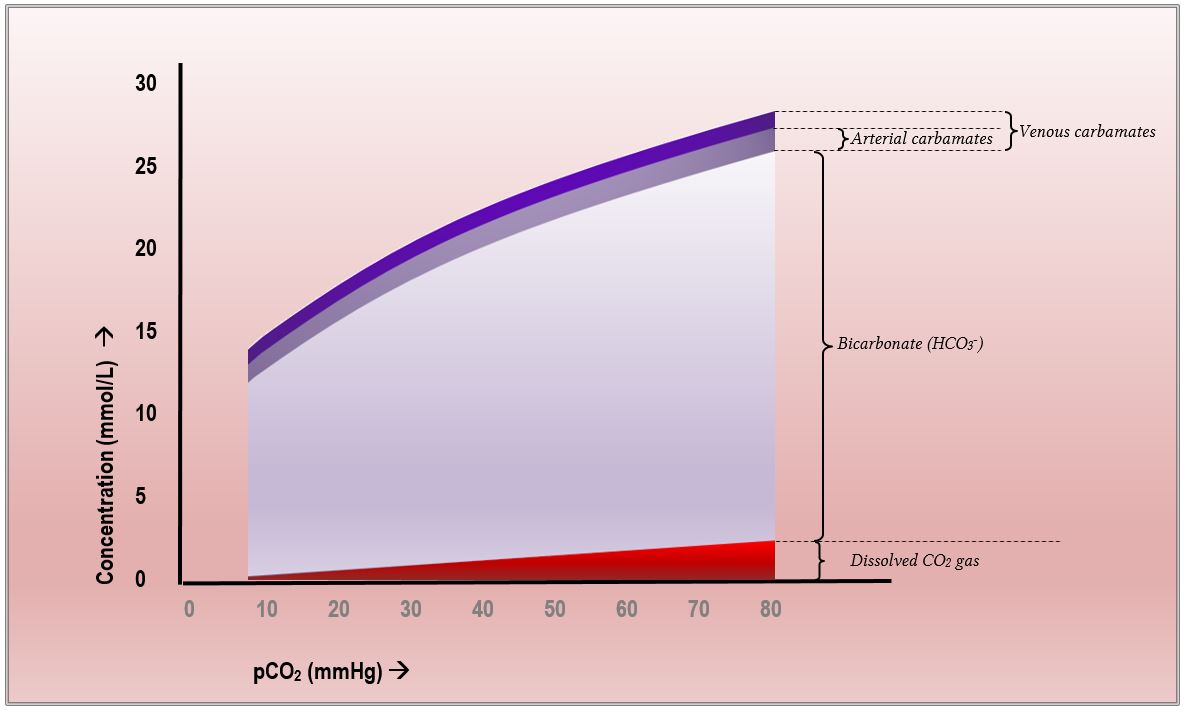
Il s’agit en fait de la courbe de dissociation du CO2, similaire à la courbe de dissociation oxygène-hémoglobine mais distincte car contrairement à l’oxygène, le CO2 est transporté de trois manières différentes. Comme on peut le voir clairement, avec l’augmentation du pCO2, la concentration de bicarbonate change le plus, le CO2 dissous augmente modestement de manière linéaire et la teneur en carbamate change à peine du tout.
transport du CO2 sous forme de gaz dissous
le CO2 se trouve être l’un des gaz qui jouissent d’une relativement bonne solubilité dans l’eau., Pour chaque 1 mmHg DE CO2, environ 0,03 mmol/L pénètre dans la solution. Cette relation est raisonnablement linéaire sur une plage de survie de pressions et de températures.

en solution, le CO2 se combine de manière réversible avec l’eau pour former de l’acide carbonique, mais il le fait avec une grande réticence. On peut décrire cette réticence mathématiquement, et l’exprimer comme une constante d’équilibre d’hydratation (constante de Henry). Cette constante est dérivée en divisant la concentration de H2CO3 par la concentration de CO2 aqueux. Ainsi, à 25°C, la constante de Henry pour le CO2 n’est que d’environ 1.,7 x10-3, ce qui signifie que dans un pot d’eau donné, la majorité du CO2 dissous reste sous forme de gaz dissous, et seule une infime fraction (0,17%) est présente sous forme de H2CO3.
de toute façon. La teneur totale en CO2 dissous dans le sang peut être calculée à partir de la relation précitée ( 0,03 mmol/l/mmHg); elle est donc de 1,2 mmol/l.
transport du CO2 sous forme de bicarbonate
Conversion du CO2 en acide carbonique et en bicarbonate
la combinaison du CO2 et du H2O conduit à la formation du H2CO3 de manière totalement spontanée non catalysée., Le CO2 agit comme un acide de Lewis dans ce contexte; ne pas avoir de protons pour le donner agit plutôt comme « quelque chose capable de former une liaison covalente avec une paire d’électrons ».
la réaction spontanée avec l’eau est en fait plutôt lente. Pour des quantités cliniquement significatives de CO2, la réaction de dissociation Non catalysée CO2 + H2O ⇌ H2CO3 peut prendre environ 3-4 secondes à demi-équilibre, et jusqu’à une minute pour atteindre l’équilibre complet (et c’est dans la température corporelle de soutien de réaction torride, 37°C). Évidemment, c’est beaucoup trop lent., Il n’y aurait pas assez de temps pour que cette réaction fonctionne à l’équilibre pendant le transit rapide des globules rouges à travers les capillaires pulmonaires échangeurs de gaz, et l’élimination du CO2 ne se produirait pas.

heureusement, l’intérieur des érythrocytes (ainsi que les parois des vaisseaux sanguins) sont remplis d’anhydrase carbonique (en particulier de type CA II) , qui est peut-être l’enzyme agissant le plus rapidement dans le corps humain, et à l’aide de laquelle cette réaction progresse 25 000 fois plus rapidement, atteignant 95% , Dans les macro-échelles de ventilation, mesurées en secondes et en minutes, cette réaction peut être considérée comme essentiellement instantanée. En raison de ce type de support catalytique, la conversion de HCO3 – et H+ en CO2 et H2O peut se produire dans les capillaires pulmonaires pendant le temps de transit capillaire pulmonaire très bref.
en fait, l’anhydrase carbonique est si efficace que relativement peu de molécules enzymatiques sont nécessaires pour maintenir un niveau d’échange de CO2 tout à fait normal. Il semblerait que l’on nécessite peut-être aussi peu que 2% d’enzyme non bloquée pour continuer., Considérer que ce système enzymatique est systématiquement bloqué pour des raisons thérapeutiques par des médicaments tels que l’acétazolamide. En effet, les surdoses massives d’inhibiteurs de l’anhydrase carbonique ont été étonnamment survivables, malgré l’acidose métabolique sévère qui s’ensuit parfois.
transport du CO2 sous forme de composés carbamates
Les Composés carbamates sont les bases conjuguées dissociées des carbamino-acides, qui se forment dans la réaction spontanée de R-NH2 et de CO2.,

ainsi, un « carbamate » est une base conjuguée qui agit comme un dispositif de stockage de CO2, et peut contribuer jusqu’à 14% de la capacité de charge totale de CO2 du sang. Le stade transitoire « carbamino » décrit ci – dessus est – comme l’acide carbonique-une figure de la parole plutôt qu’un membre macroscopique approprié de la circulation sanguine., Au pH physiologique, il se dissocie en carbamate et en h+ si rapidement et si complètement que sa concentration est négligeable, et à toutes fins utiles, il peut être
la teneur en carbamate du plasma est plutôt médiocre car les protéines sériques (par exemple. globulines) ont peu d’intérêt pour le transport du CO2, et ont tendance à former des carbamates avec la plus grande réticence. Le CO2 ne peut vraiment se lier qu’aux chaînes latérales de la lysine et de l’arginine, et toute protéine plasmatique aléatoire ne peut avoir qu’une de ces chaînes latérales exposée et disponible pour la liaison au CO2., Plus de sites de liaison sont disponibles dans les globules rouges, où l’hémoglobine (en particulier la désoxyhémoglobine) est disponible à une concentration élevée.
cette rareté des sites de liaison pourrait faire penser que les carbamates ne jouent pas beaucoup de rôle dans le transport du CO2 en général, et en particulier son transport dans le plasma. En effet, les carbamates contribuent relativement peu à la capacité globale de CO2 du sang humain. Leur importance devient beaucoup plus grande si, pour une raison quelconque, la majeure partie de votre anhydrase carbonique est désactivée., Dans ce cas, ce système de transport oublié devient soudainement très pertinent, car il peut fonctionner avec du CO2 brutet ne nécessite pas de H2CO3 comme intermédiaire. De plus, le système carbamate apporte la plus grande contribution à la différence entre la concentration artérielle et veineuse de CO2, principalement en raison de l’effet Haldane qui est discuté ailleurs.,
facteurs influençant la capacité de charge en CO2 des protéines
le transport du CO2 sous forme de carbamates est influencé par les facteurs suivants:
- oxygénation de l’hémoglobine (plus désoxygénée= plus grande capacité en CO2)
- pH dans l’environnement local (pH inférieur = plus grande capacité en CO2)
à l’intérieur des globules rouges, le CO2 se lie facilement aux chaînes latérales des acides aminés de la molécule d’hémoglobine, en particulier lorsqu’elle est désoxygénée. En effet la désoxyhaémoglobine en contient environ 3.,5 fois l’affinité pour le CO2 par rapport à l’oxyhaémoglobine; cette augmentation induite par l’hypoxie de la capacité de charge du CO2 est appelée L’effet Haldane. C’est exactement ce que vous voulez dans l’exercice du tissu, qui vient de consommer tout l’oxygène dans son volume sanguin local, et a produit une grande quantité de CO2. Ainsi, la teneur en carbamate du sang veineux pourrait potentiellement (à un SvO2 de 0%) être environ le triple de la teneur en carbamate du sang artériel. Bien sûr, en réalité, la différence a-v n’est jamais aussi grande, car le sang veineux n’est jamais aussi hypoxique.,
la présence de groupes carbamates dans les régions critiques de la molécule d’hémoglobine tend à stabiliser la forme désoxygénée, ce qui diminue l’affinité de liaison hémoglobine-oxygène. Cet effet (L’effet Bohr) est discuté plus en détail avec les autres facteurs qui affectent l’affinité de l’hémoglobine pour l’oxygène. Il suffit de dire que la présence d’une grande quantité de CO2 diminue cette affinité et favorise la libération de plus d’oxygène de l’hémoglobine, ce qui est précisément ce qui est nécessaire dans les tissus d’exercice.




