La LTP a été étudiée de manière approfondie dans l’hippocampe des mammifères, une zone du cerveau qui est particulièrement importante dans la formation et/ou la récupération de certaines formes de mémoire (voir Chapitre 31). Chez l’homme, l’imagerie fonctionnelle montre que l’homme hippocampe est activée lors de certains types de tâches de mémoire, et que les dommages à l’hippocampe, entraîne une incapacité à former certains types de nouveaux souvenirs. Chez les rongeurs, les neurones de l’hippocampe ne déclenchent des potentiels d’action que lorsqu’un animal se trouve à certains endroits., De telles « cellules de lieu » semblent coder des mémoires spatiales, une interprétation étayée par le fait que les dommages de l’hippocampe empêchent les rats de développer des compétences dans les tâches d’apprentissage spatial (Figure 25.4). Bien que de nombreuses autres zones du cerveau soient impliquées dans le processus complexe de formation, de stockage et de récupération de la mémoire, ces observations ont conduit de nombreux chercheurs à étudier cette forme particulière de plasticité synaptique dans l’hippocampe.

Figure 25.4
l’apprentissage Spatial chez les rongeurs., A) les Rats sont placés dans une arène circulaire (de la taille et de la forme d’une pataugeoire pour enfants) remplie d’eau trouble. L’arène elle-même est sans caractéristiques, mais l’environnement environnant contient les indices de position tels que les fenêtres, (plus…
Les travaux sur la LTP ont commencé au début des années 1970, lorsque Timothy Bliss et ses collègues de Mill Hill en Angleterre ont découvert que quelques secondes de stimulation électrique à haute fréquence peuvent améliorer la transmission synaptique dans l’hippocampe du lapin pendant des jours, voire des semaines., Plus récemment, cependant, les progrès dans la compréhension du mécanisme de la LTP se sont largement appuyés sur des études in vitro sur des tranches d’hippocampe vivant. La disposition des neurones permet à l’hippocampe d’être sectionné de telle sorte que la plupart des circuits pertinents soient laissés intacts. Dans de telles préparations, les corps cellulaires des neurones pyramidaux se trouvent dans une seule couche densément compacte qui est facilement apparente (Figure 25.5). Cette couche est divisée en plusieurs régions distinctes, les principales étant CA1 et CA3., « CA » fait référence à Cornu Ammon, un nom Latin pour la corne D’Ammon—la corne du bélier qui ressemble à la forme de l’hippocampe. Les dendrites des cellules pyramidales dans la région CA1 forment une bande épaisse (la strate radiatum), où elles reçoivent des synapses des collatéraux de Schaffer, les axones des cellules pyramidales dans la région CA3. Une grande partie du travail sur la LTP a porté sur les connexions synaptiques entre les collatéraux de Schaffer et les cellules pyramidales CA1. La stimulation électrique des collatéraux de Schaffer génère des potentiels postsynaptiques excitateurs (EPSPs) dans les cellules CA1 postsynaptiques (Figure 25.6)., Si les collatéraux de Schaffer ne sont stimulés que deux ou trois fois par minute, la taille de L’EPSP évoqué dans les neurones CA1 reste constante. Cependant, un bref train de stimuli à haute fréquence vers les mêmes axones provoque une LTP, ce qui est évident comme une augmentation durable de L’amplitude EPSP. La LTP se produit également à de nombreuses autres synapses, à la fois dans l’hippocampe et dans une variété d’autres régions du cerveau, y compris le cortex, l’amygdale et le cervelet.

la Figure 25.,5
schéma d’une section à travers l’hippocampe du rongeur montrant les principales régions, les voies excitatrices et les connexions synaptiques. Une potentialisation à Long terme a été observée à chacune des trois connexions synaptiques présentées ici.
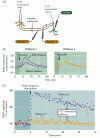
Figure 25.6
potentialisation à Long terme des synapses collatérales de Schaffer-CA1. (A) agencement pour enregistrer la transmission synaptique; deux électrodes stimulantes (1 et 2) activent chacune des populations distinctes de garanties Schaffer, fournissant ainsi un test et un contrôle synaptique (more…,)
LTP de la synapse collatérale de Schaffer présente plusieurs propriétés qui en font un mécanisme neuronal attrayant pour le stockage d’informations. Premièrement, la LTP dépend de l’état: le degré de dépolarisation de la cellule postsynaptique détermine si la LTP se produit ou non (Figure 25.7). Si un seul stimulus des collatéraux de Schaffer – qui ne déclencherait normalement pas de LTP-est associé à une forte dépolarisation de la cellule CA1 postsynaptique, la taille de L’EPSP est augmentée., L’augmentation ne se produit que si les activités appariées des cellules présynaptiques et postsynaptiques sont étroitement liées dans le temps, de sorte que la forte dépolarisation postsynaptique se produit dans environ 100 ms de libération de l’émetteur présynaptique. Rappelons qu’une exigence d’activation concomitante d’éléments présynaptiques et postsynaptiques est la caractéristique du postulat Hebbian (voir Chapitre 24), une première tentative d’établir un cadre théorique des changements synaptiques sous-jacents à l’apprentissage et à la mémoire.

la Figure 25.,7
L’appariement de l’activité présynaptique et postsynaptique provoque une LTP. Des stimuli simples appliqués à une entrée synaptique collatérale de Schaffer évoquent des EPSP dans le neurone CA1 postsynaptique. Ces stimuli seuls ne provoquent aucun changement dans la force synaptique. Cependant, lorsque le (plus…
La LTP présente également la propriété de spécificité d’entrée: lorsque la LTP est induite par la stimulation d’une synapse, elle ne se produit pas dans d’autres synapses inactives qui entrent en contact avec le même neurone (voir Figure 25.6)., Ainsi, la LTP est spécifique à l’entrée en ce sens qu’elle est limitée aux synapses activées plutôt qu’à toutes les synapses d’une cellule donnée (Figure 25.8 A). Cette caractéristique de LTP est compatible avec son implication dans la formation de la mémoire. Si l’activation d’un ensemble de synapses entraînait la potentialisation de toutes les autres synapses—même inactives—, il serait difficile d’améliorer sélectivement certains ensembles d’entrées, comme cela est vraisemblablement nécessaire pour l’apprentissage et la mémoire (voir également L’encadré B).
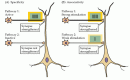
la Figure 25.,8
les Propriétés de la LTP à un pyramidaux CA1 neurone qui reçoit des entrées synaptiques à partir de deux ensembles indépendants de collatérale de Schaffer axones. (A) une forte activité initie la LTP aux synapses actives (voie 1) sans initier la LTP aux synapses inactives voisines (voie (plus…)
Zone B
Synapses Silencieuses.
Une autre propriété importante de LTP est l’associativité (Figure 25.8 B). Comme indiqué, une faible stimulation d’une voie ne déclenchera pas à elle seule la LTP., Cependant, si une voie est faiblement activée en même temps qu’une voie voisine sur la même cellule est fortement activée, les deux voies synaptiques subissent une LTP. Cette amélioration sélective d’ensembles d’entrées synaptiques activés conjointement est souvent considérée comme un analogue cellulaire du conditionnement associatif ou classique. Plus généralement, l’associativité est attendue dans tout réseau de neurones qui relie un ensemble d’informations à un autre.,
bien qu’il existe clairement un écart entre la compréhension de la LTP des synapses de l’hippocampe et la compréhension de la plasticité comportementale, cette forme de plasticité synaptique fournit un mécanisme neural plausible pour des changements durables dans une partie du cerveau connue pour être impliquée dans la formation de certains types de souvenirs.




