LTP se ha estudiado más a fondo en el hipocampo de mamíferos, un área del cerebro que es especialmente importante en la formación y/o recuperación de algunas formas de memoria (ver Capítulo 31). En los humanos, las imágenes funcionales muestran que el hipocampo humano se activa durante ciertos tipos de tareas de memoria, y que el daño al hipocampo resulta en una incapacidad para formar ciertos tipos de nuevos recuerdos. En los roedores, las neuronas del hipocampo disparan potenciales de acción solo cuando un animal está en ciertos lugares., Tales «células de lugar» parecen codificar memorias espaciales, una interpretación apoyada por el hecho de que el daño del hipocampo impide que las ratas desarrollen competencia en tareas de aprendizaje espacial (figura 25.4). Aunque muchas otras áreas del cerebro están involucradas en el complejo proceso de formación, almacenamiento y recuperación de la memoria, estas observaciones han llevado a muchos investigadores a estudiar esta forma particular de plasticidad sináptica en el hipocampo.

Figura 25.4
el aprendizaje Espacial en los roedores., (A) las ratas se colocan en una arena circular (aproximadamente del tamaño y la forma de la piscina infantil) llena de agua turbia. La arena en sí no tiene rasgos distintivos, pero el entorno circundante contiene las señales posicionales como ventanas, (más…)
El Trabajo en LTP comenzó a principios de la década de 1970, cuando Timothy Bliss y sus colegas de Mill Hill en Inglaterra descubrieron que unos segundos de estimulación eléctrica de alta frecuencia pueden mejorar la transmisión sináptica en el hipocampo de conejo durante días o incluso semanas., Más recientemente, sin embargo, el progreso en la comprensión del mecanismo de la LTP se ha basado en gran medida en estudios in vitro de cortes de hipocampo vivo. La disposición de las neuronas permite que el hipocampo sea seccionado de tal manera que la mayoría de los circuitos relevantes se dejan intactos. En tales preparaciones, los cuerpos celulares de las neuronas piramidales se encuentran en una sola capa densamente empaquetada que es fácilmente aparente (figura 25.5). Esta capa se divide en varias regiones distintas, las principales son CA1 y CA3., «CA» se refiere a Cornu Ammon, un nombre latino para el cuerno de Ammon – el cuerno de carnero que se asemeja a la forma del hipocampo. Las dendritas de las células piramidales en la región CA1 forman una banda gruesa (el estrato radiato), donde reciben sinapsis de los colaterales de Schaffer, los axones de las células piramidales en la región CA3. Gran parte del trabajo en LTP se ha centrado en las conexiones sinápticas entre los colaterales de Schaffer y las células piramidales CA1. La estimulación eléctrica de los colaterales de Schaffer genera potenciales postsinápticos excitatorios (EPSP) en las células CA1 postsinápticas (figura 25.6)., Si los colaterales de Schaffer son estimulados solo dos o tres veces por minuto, el tamaño de la EPSP evocada en las neuronas CA1 permanece constante. Sin embargo, un breve tren de estímulos de alta frecuencia a los mismos axones causa LTP, lo que es evidente como un aumento duradero en la amplitud de la EPSP. La LTP también ocurre en muchas otras sinapsis, tanto dentro del hipocampo como en una variedad de otras regiones del cerebro, incluyendo la corteza, la amígdala y el cerebelo.

Figura 25.,5
diagrama de una sección a través del hipocampo del roedor que muestra las principales regiones, vías excitatorias y conexiones sinápticas. Se ha observado una potenciación a largo plazo en cada una de las tres conexiones sinápticas que se muestran aquí.
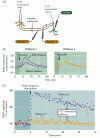
Figura 25.6
la potenciación a Largo plazo de colateral de Schaffer-CA1 sinapsis. (A) disposición para registrar la transmisión sináptica; dos electrodos estimulantes (1 y 2) activan cada uno poblaciones separadas de colaterales de Schaffer, proporcionando así Prueba y control sináptico (más…,)
LTP de la sinapsis colateral de Schaffer exhibe varias propiedades que la convierten en un mecanismo neuronal atractivo para el almacenamiento de información. En primer lugar, la LTP es dependiente del Estado: el grado de despolarización de la célula postsináptica determina si se produce o no LTP (figura 25.7). Si un único estímulo a los colaterales de Schaffer-que normalmente no provocaría LTP-se aparea con una fuerte despolarización de la célula postsináptica CA1, el tamaño de la EPSP aumenta., El aumento ocurre solo si las actividades pareadas de las células presinápticas y postinápticas están estrechamente vinculadas en el tiempo, de modo que la fuerte despolarización postináptica ocurre dentro de unos 100 ms de la liberación del transmisor presináptico. Recordemos que un requisito para la activación coincidente de elementos presinápticos y postsinápticos es el sello distintivo del postulado Hebbiano (ver capítulo 24), un intento temprano de establecer un marco teórico de los cambios sinápticos subyacentes al aprendizaje y la memoria.

Figura 25.,7
El emparejamiento de actividad presináptica y postsináptica causa LTP. Los estímulos individuales aplicados a una entrada sináptica colateral de Schaffer evocan EPSPs en la neurona CA1 postsináptica. Estos estímulos por sí solos no provocan ningún cambio en la fuerza sináptica. Sin embargo, cuando el (más…)
la LTP también exhibe la propiedad de la especificidad de entrada: cuando la LTP es inducida por la estimulación de una sinapsis, no ocurre en otras sinapsis inactivas que entran en contacto con la misma neurona (ver figura 25.6)., Por lo tanto, LTP es específico de entrada en el sentido de que está restringido a sinapsis activadas en lugar de a todas las sinapsis en una celda dada (figura 25.8 A). Esta característica de LTP es consistente con su participación en la formación de la memoria. Si la activación de un conjunto de sinapsis llevó a todas las otras sinapsis—incluso las inactivas—potenciadas, sería difícil mejorar selectivamente conjuntos particulares de entradas, como se supone que se requiere para el aprendizaje y la memoria (Véase también el recuadro B).
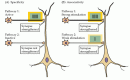
Figura 25.,8
propiedades de LTP en una neurona piramidal CA1 que recibe entradas sinápticas de dos conjuntos independientes de axones colaterales de Schaffer. (A) la actividad Fuerte inicia LTP en sinapsis activas (vía 1) sin iniciar LTP en sinapsis inactivas cercanas (vía (más…)
Caja B
Silencio de las Sinapsis.
otra propiedad importante de LTP es la asociatividad (figura 25.8 B). Como se señaló, la estimulación débil de una vía no desencadenará LTP por sí sola., Sin embargo, si una vía se activa débilmente al mismo tiempo que una vía vecina a la misma célula se activa fuertemente, ambas vías sinápticas experimentan LTP. Esta mejora selectiva de conjuntos activados conjuntamente de entradas sinápticas a menudo se considera un análogo celular del condicionamiento asociativo o clásico. Más generalmente, se espera asociatividad en cualquier red de neuronas que vincule un conjunto de información con otra.,
aunque existe claramente una brecha entre la comprensión de la LTP de las sinapsis del hipocampo y la comprensión de la plasticidad conductual, esta forma de plasticidad sináptica proporciona un mecanismo neural plausible para cambios duraderos en una parte del cerebro que se sabe que está involucrada en la formación de ciertos tipos de recuerdos.




